Information station of Parasitoid wasps
Information station of Parasitoid wasps

ヒメバチ科の形態
以下に、図を元にヒメバチの形態について解説を加えてゆきます。もちろん、コマユバチなど、他のハチにも多くは適用できます。各形態についての説明は併せて形態用語辞典を参照ください。
なお、いくつかの図は、多くの形質を説明する為、実際の種(標本)を用いていないものであることに注意して下さい。
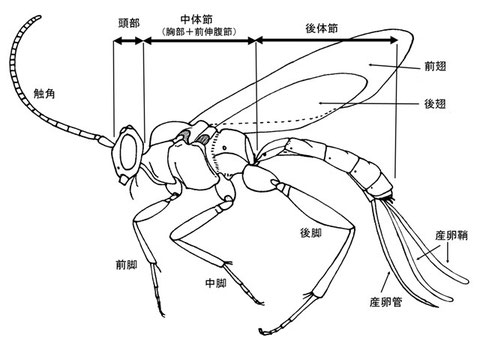
大まかな形態的特徴
ハチの体も他の昆虫同様、頭部(Head)、胸部(Thorax)、腹部(Abdomen)の3つの部分にわけられるが、細腰亜目のハチでは、真の腹部第1節が前伸腹節(Propodeum)という部分として胸部と融合する。その為、外見上の腹部は真の腹部第2節以降ということになる。そこで、混乱を防ぐ為に、ハチの研究者間では外見上の胸部を中体節(Mesosoma)、外見上の腹部を後体節(Metasoma)として扱う。
胸部もまた他の多くの昆虫同様、3つの部分(前胸・中胸・後胸)にわかれ、中胸からは前翅(Forewing)、後胸からは後翅(Hindwing)、そして各節より1対の脚が生えている。ヒメバチを含む多くの寄生蜂では、転節と腿節の間に、第二転節という部分を持つ(詳しくは脚の解説を参照)。
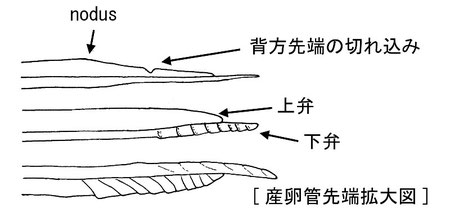
しばしば、ヒメバチには産卵管が3本あるのか?という質問をうけますが、うち2本は、産卵管を保護する産卵鞘です。よく見てみると、構造がまったく違うことに気がつくはずです。
方向に関しては、以下のような用語を用います。
日本語での意味 大凡該当する用語
前方 Anterior, Front, Frontal
後方 Posterior, Rear, Back
背方 Dorsal
腹方 Ventral
上方 Above
下方 Below
中央 Median, Medial, Middle, Mid-
側方 Lateral
先端方 Apical, Distal
先端 Apex
基部方 Basal, Proximal
基部 Base
頭部 HEAD
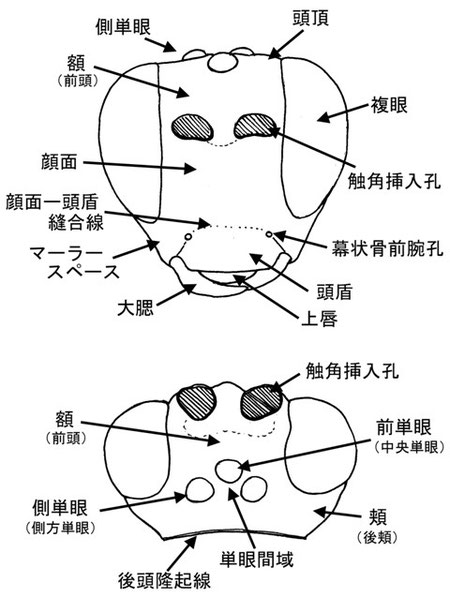
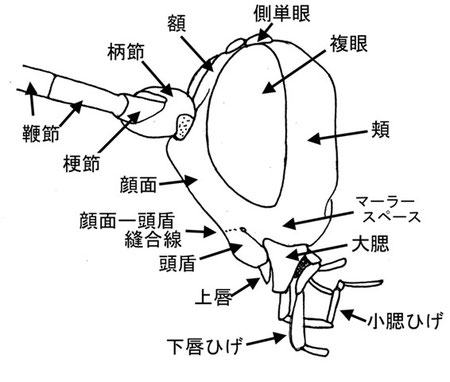
頭部の主要な部位を上記に図示する。境界については時に不明瞭で、頬と顔面あるいは頭頂の境界は大抵の場合わかりにくい。顔面と額(前頭)の境界は両方の触角挿入孔の下縁を結ぶ水平線を境界とし、額と頭頂の境界は中央単眼前縁における同様な水平線である。顔面と頭盾の境界は前頭‐頭盾縫合線であり、これは幕状骨前腕孔、大腮基部のそれぞれの間に存在するが、しばしば消失する。この場合は、各幕状骨前腕孔間を結ぶ水平線を仮定し、そこを境界として扱う。計測箇所は研究者によって様々であるので、その都度確認する必要がある。
マーラースペースは頬の一部であり、その長さは複眼と大腮間の長さを図る重要な目安である。その距離は複眼下縁と大腮基部の内方を最短距離で結んだ距離である。通常はその長さを大腮基部の幅と比較する。
大腮の先端は通常2歯を持つが、しばしば1歯になったり、二次的に分割し3歯になる。また、大腮は基部に窪みを有したり、隆起を有したり、ねじれたりもする。
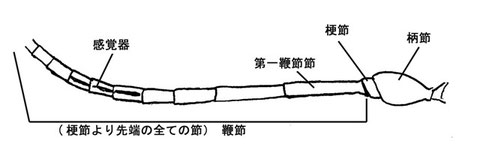
触角は寄主探索などに強く関係する為、グループによってはしばしば顕著な特徴があらわれることがある。例えばフトマルヒメバチ亜科の♂は中央が拡大し、ホソヒメバチ亜科(Cylloceria)の♂では鞭節の基部方の節に三日月型の切れ込みがある。また、トガリヒメバチ亜科(Thyribiusなど)の一部などでは先端が裁断状のときもあり、マルズヒメバチ亜科(Xorides)やウスマルヒメバチ亜科(Banchus)では著しい感覚毛を先端方に有する。さらに、通常確認される小さな感覚器以外にヒラタヒメバチ亜科やヒラタアブヤドリヒメバチ亜科の一部等では瘤状の著しい感覚器を有することがあり、感覚器という用語は通常これに対し用いる。
鞭節の節数、長さは分類においてしばしばとても重要である。
中体節 MESOSOMA
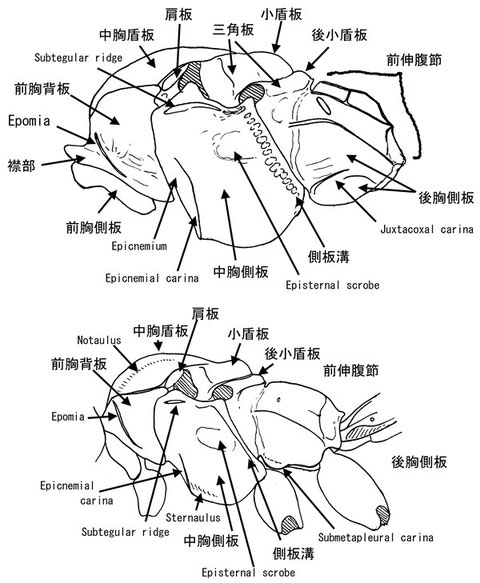
中体節を側方から見た際の主要な形質を上記に図示する。前胸背板は側方後方へと発達し、多くのグループでEpomiaと称される隆起線を有する。この隆起線の後方はしばしば広い無点刻域や、強い皺に覆われることがあり、それらは種の識別に時に有用である。中胸背板は外見上中胸盾板と小盾板に分けて扱われ、両者間は溝で区分される。前者は大抵、中胸背縦斜溝(Notauli)を持ち、これらの発達度合いは分類に有用である。後者はしばしば側方に隆起線を伴うことがある。また、一部のグループでは先端に棘や瘤をもつ。中胸側板は重要な部位で、Epicnemial carinaや胸側溝、側板溝、Subalar ridgeの発達度合いは分類、とくに高次分類において重要である。また、無点刻域や皺に覆われた領域の面積は種分類に有用な事が多い。後胸側板は一部のグループで表面彫刻が分類に用いられる。前伸腹節は隆起線によって様々な区画に分割される。隆起線は部分的に減少することが多々あり、それに伴い区画間で融合が生じることがある。前伸腹節の気門はしばしば伸長することがあり、形は高次分類に用いられることがある。
翅 WINGS
翅は前翅と後翅、それぞれ中胸と後胸に1対ずつ存在するが、一部のグループでは短翅型や無翅型が存在する。
翅脈ならびに翅室の名称は、研究者によって意見が相違することもあり、いくつかの体系があるが、代表的なものに、Gauld (1991)とTownes (1969)がある。後者はやや古いが、特に前翅のNervulus と 後翅のNervellus について便利であり、現在でも使用している研究者がいる。
なお、Hymenoptera of the Worldで使用されている体系はコマユバチの形態の頁を参照いただきたい。
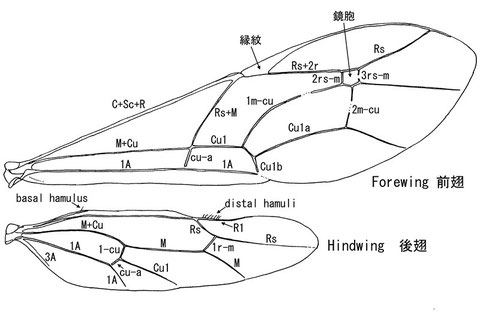
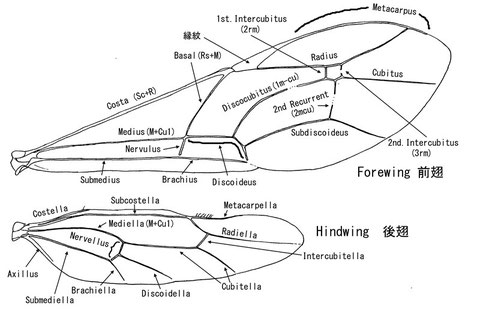
翅脈の計測は、接合点の中心と中心を結んで計測する。各翅脈にはそれぞれ日本語の訳があるが、翅脈に関して言えばそのまま表記した方がはるかに便利である。
たくさんの翅脈のうち、良く出てくる箇所は大まかに3箇所であり、鏡胞とその周辺、前翅翅脈Rs+M脈とcu-a脈のそれぞれの起点の位置関係、そして後翅翅脈1-cuとcu-aの長さ、傾き、分岐する場合のCu1の存在、である。以下に模式図を用いて代表的なパターンについて説明を行う(図の左が基部方、右が先端方であり、右翅を用いて説明しているとイメージしていただきたい)。
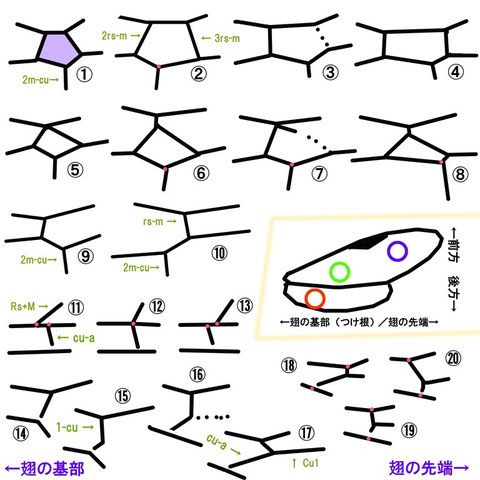
鏡胞(図①の紫色に塗った箇所)はその有無、形のほか、2m-cu脈を受け入れる位置がよく利用される。
図①~⑩は紫色の丸で示したあたりにある鏡胞と周囲を示している。以下に状態と該当する図を示す。
・鏡胞は存在する(鏡胞は閉じる)・・・①~⑧
・鏡胞は欠く(鏡胞は開く)・・・⑨、⑩
ただし、研究者によっては開くの形質状態に③をあてることもある
・鏡胞は五角形・・・①~④
・鏡胞は四角形・・・⑤~⑧(⑤と⑥はひし形)
・鏡胞の前方は尖る・・・⑤~⑧
・鏡胞の前方は柄をもつ(有柄)・・・⑥、⑧(短い柄)
・2m-cu脈は鏡胞の基部よりから生じる・・・②、⑦
・2m-cu脈は鏡胞の中央付近よりから生じる・・・⑤、⑥
・2m-cu脈は鏡胞の先端よりから生じる・・・①(やや先端)、③、④、⑧
・2m-cu脈は鏡胞の外角付近から生じる・・・⑧
・2m-cu脈はrs-m脈(3rs-m脈が無い場合は2rs-mといわずrs-mという)より翅の先端方から生じる・・・①~⑨
・2m-cu脈はrs-m脈より翅の基部方から生じる・・・⑩(コンボウアメバチ亜科やアメバチ亜科などで見られる)
・鏡胞の側縁は平行(2rs-m脈と3rs脈は平行)・・・④
・鏡胞の側縁は前方に向かって幅が狭くなる・・・①~③、⑤~⑨
これら表現はトガリヒメバチ亜科の同定でしばしば使う。
前翅翅脈cu-a(Nervulus) はRs+M(Basal vein) との位置関係でよく使われ、双方の起点が比較される。以下に状態と該当する図を示す。
図⑪~⑬は黄緑色の丸で示したあたりを示している。
・cu-aの起点はRs+Mの起点よりも先端よりに存在する(=翅脈自体の幅の2倍程度先端よりから生じる)・・・⑪
・cu-aの起点はRs+Mの起点と対の位置に存在する・・・⑫
・cu-aの起点はRs+Mの起点よりも基部よりに存在する・・・⑬
後翅翅脈1-cuとcu-a(Nervellus)はどこで折れるかが使われ、たいていは折れた箇所から翅脈Cu1(Discoidella)が生じる。そしてその傾きも重要である。
図⑭~⑳はオレンジ色の丸で示したあたりを示している。
・折れる・・・⑮~⑳
・前方で折れる・・・⑰、⑱
・中央付近で折れる・・・⑯、⑲
・後方で折れる・・・⑮、⑳
・折れない・・・⑭
ちなみに、Nervellusは前方0.3付近で折れる、という場合は1-cuとcu-aを足した全体の長さを10分割し、前方から3区画目付近で折れることをいい。⑰のようなケースをいう。
・Cu1を欠く・・・⑭、⑮
・Cu1は不明瞭(痕跡状、光の加減で)に存在する・・・⑯
・Cu1を有する(明瞭)・・・⑰~⑳
・Reclivous(外斜):横脈の前方と後方の終点を見たとき、後方終点の方が基部に近いこと・・・⑰、⑱(⑭~⑯も翅の付け根の位置によっては該当)
・Vertical (垂直):横脈の前方と後方、双方の終点が基部と(ほぼ)等しい位置にあること・・・⑲(⑭~⑯も翅の付け根の位置によっては該当)
・Inclivous(内斜):横脈の前方と後方の終点を見たとき、前方終点の方が基部に近いこと・・・⑳
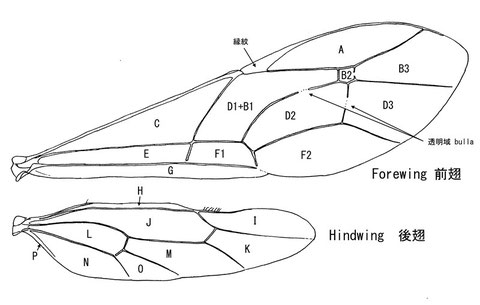
翅室の名称
(Gauld, 1991の体系)
A & I: Marginal cell
D1+B1: Discosubmarginal cell
B2: Areolet
C & J: Basal cell
D2: 2nd. discal cell
E & L: subbasal cell
F1: 1st. subdiscal cell
(Townes, 1969の体系)
A: Radial cell
B: Cubital cell
D1+B1: Discocubital cell
B2: Areolet
B3: 3rd. cubital cell
C: Median cell
D: Discoidal cell
D2: 2nd. discoidal cell
D3: 3rd: discoidal cell
E: Submedian cell
F1: 1st. brachial cell
F2: 2nd. brachial cell
G: Anal cell
H: Costellan cell
I: Radiellan cell
J: Mediellan cell
K: Cubitellan cell
L: Submediellan cell
M: Discoidellan cell
N: Anellan cell
O: Brachiellan cell
P: Postellan cell
脚 LEG
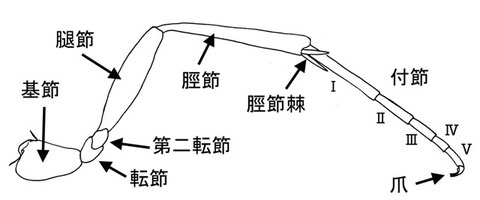
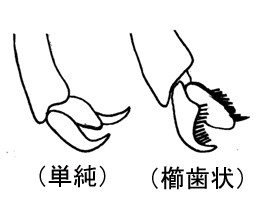
脚の形質を、左脚の後脚を側方から見た図で説明する。ここではtrochantellusに対し、第二転節という名称を与えている。これは、文献によってもちいられている小転節という名称が他の部位にも用いられている為である。この第二転節は、転節とは異なり、腿節の一部がから生じたとされている。脛節棘の本数はしばしば減少し、前脚においては常に1本で、触角の清掃などに使用する。爪は厳密には前付節に生じるが、日本語名称では付節爪として利用する。爪はしばしば付属歯や櫛歯状の歯を持つことがある。
脚はしばしば特殊化した形質を持つが、それらの生態的な役割については殆ど論じられていない。
後体節 METASOMA
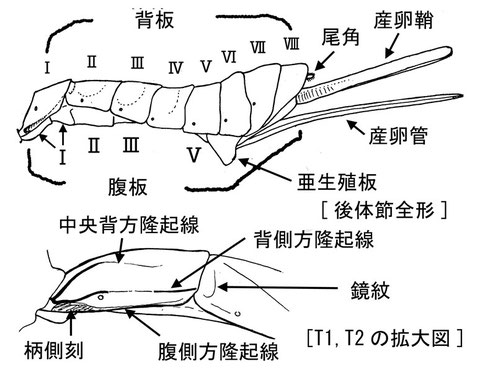
後体節の各節は、実際の腹部の節と1節づつずれているが、便宜を考え、一般に第1背板というときは「後体節の」第1背板を指す。背板(Tergite)、腹板(Sternite)はしばしば、それぞれの頭文字をとってT1, T2..., S1, S2...と略記されることがある。後体節第1背板の気門の位置は、高次分類においてきわめて重要である。後体節背板の長さと幅は分類に有用であるが、その計測基準は論文によって様々である。
産卵管はその構造と長さが分類においてきわめて重要である。なぜならば各種が寄主に適応する際、最もその特徴が関係する部位のためである。利用する(できる)寄主の違いで種の分化が生じているためか、一般に雄交尾器の形態差は少ない。ただし、夜行性のアメバチモドキ(Netelia)などでは、雄交尾器の形態が種間で明瞭に異なることもある。産卵管の長さは一般的に体外に露出している部分の長さで計測され、それは産卵鞘の長さと一致すると(厳密にはしないかもしれないが)して、産卵管露出部か産卵鞘のいずれかの長さで計測される。もし前者を計測する場合、産卵鞘の付け根までの長さを差し引いて計測すればよい。計測した長さは、一般に後脚脛節、後脚付節、前翅などと比較されるが、前翅は標本のコンディションによってはゆがむために比較が難しい。